

➤ Steve Leeson
Professor Emeritus, University of Guelph
sleeson@uoguelph.ca
Introduction
Energy is the most costly component of poultry diets, with global spending exceeding $100 billion. Unlike other nutrients, there is little assurance of the energy content in purchased ingredients or formulated diets, and current estimates rely on expensive and time-consuming bioassays with live birds. Alternative predictive methods, such as equations, in vitro assays, or rapid physical analyses, have had limited success.
Energy is also the main driver of feed intake: birds eat to meet their energy requirements. Therefore, setting diet energy levels directly affects the proportional levels of other nutrients, such as amino acids. White-egg layers adjust feed intake very accurately according to diet energy, while broilers can do so only under true ad libitum conditions, which are rare in commercial settings.
Accurate knowledge of diet energy is therefore essential for proper nutrient formulation. The following discussion focuses on energy evaluation systems, energy requirements, and potential alternatives for meeting birds’ energy needs.
Energy evaluation
For the last 50 years or so we have used the schematic of energy definition to describe possible options for evaluation (Figure 1).
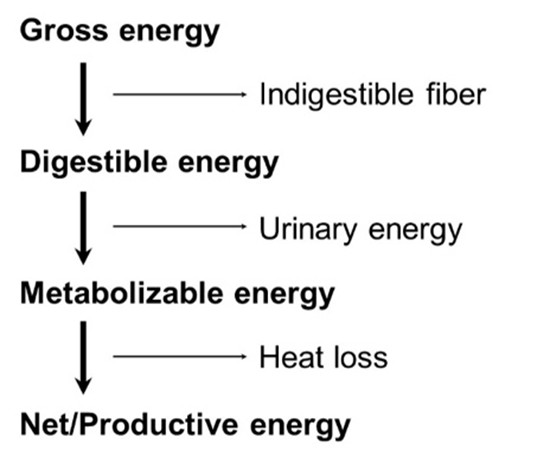
We invariably dismiss GE since it represents total heat of combustion. However, for QC programs we need to identify “outliers” and so GE can give us a rapid indication that perhaps an ingredient is different from the norm, and so warrants further investigation. DE is likewise dismissed as being impractical to measure since the bird excretes urine and faeces combined, and so measurement needs either surgical modification of the bird or sampling from the distal intestine. On the other hand, we have embraced DAAs (Digestible Amino Acids) as a standard in describing ingredients and diets that are now routinely used in formulation. It is somewhat surprising that we did not pursue DE in the same manner. DE has the major advantage over ME and NE of being totally unencumbered by bird status and general feeding status. DE is arguably the most appropriate system of evaluating and ranking individual ingredients.
Metabolizable energy has become the system of choice in evaluating ingredients and diets and virtually all recommendations from commercial breeding companies are as ME. Although we talk about ME, in fact we are usually referring to AMEn. The ’A’ prefix refers to the “Apparent” status of these values, since there are always endogenous energy losses from a bird regardless of feed intake. The industry briefly looked at TMEn that accounts for endogenous losses. Such losses are quite minor, unless the bird is given very limited quantities of feed. With ad libitum intake, endogenous losses are in the order of 2-3%, which has a minor impact in the calculation of retained energy. Such endogenous losses are inevitable and must be “paid-for” at some stage in the calculation of energy retention, a situation that occurs with AMEn. To some extent endogenous losses will vary with ingredient and is usually higher for ingredients with higher levels of fibre (so contributing to their lower energy capture).
The suffix “n” in AMEn refers to nitrogen retention status and is perhaps of more significance. During a bioassay, birds will retain nitrogen to varying degrees and so this impacts urinary energy output. The greater the nitrogen retention, the higher the AME value. To reduce such variance, the common practice is to correct all assay calculations based on the assumption that no nitrogen is retained i.e. zero nitrogen retention. Each gram of N retained, if it appeared in the urine would add 34.4 kJ to the excreta energy. This rather drastic measure has some rational basis in that all N/protein is eventually used as an energy source during protein turnover. Labelled N fed to the broiler at 7 days will not be found in the body at 42 days for example. However, during a 4-5 day bioassay, obviously, all N consumed will not be excreted in a healthy bird. The zero-retention basis also had a legitimate basis when introduced in the 1960s since nutritionists were formulating diets for multiple farm species that had inherently different rates of N retention. However, it seems somewhat illogical to derive energy values accepting the situation that the bird is N-free and has not grown, or that the laying hen has not produced eggs. One advantage of using adult roosters in such ME bioassays is that they are usually close to zero-N balance and so the correction is a physiological norm. However, we know that age of bird impacts AME values (Lopez and Leeson, 2007). The n-correction will obviously penalise protein-rich ingredients more so than cereals. Since broiler nutritionists rarely need to formulate for multi-species that perhaps have variable N-retention, then perhaps the use of uncorrected values (AME) is more appropriate for today’s more focused poultry nutrition. Lopez and Leeson (2008) estimated both AME and AMEn for maize and soybean meal with different ages of broiler (Table 1)
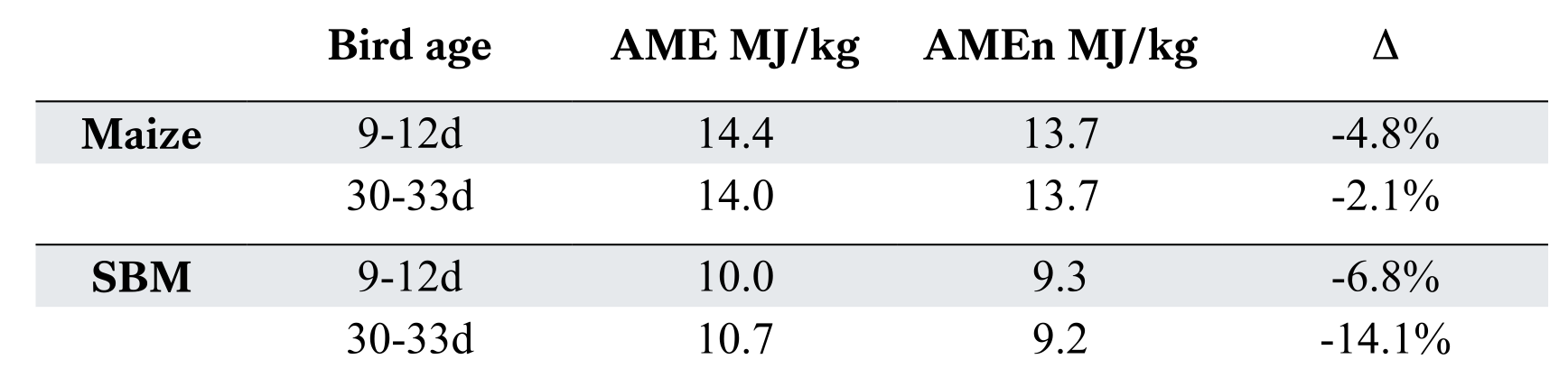
As suspected, the correction for N retention penalizes SBM by 7-14% compared to just 2-5% with maize. Abdollahi et al (2021) showed comparable differences for SBM. In taking this concept of AME for broilers one step further, Lopez and Leeson (2008) showed that broilers grown on diets formulated to AME, rather than conventional AMEn, had slightly inferior growth rate (-20g) and inferior feed efficiency (+0.02) although the cost per kg of body weight gain was reduced by 5%.
Formulating to AME does however pose some practical issues. Because N retention varies with age, then age-related AME values are likely needed to fully capitalize on this system. The other major obstacle currently, is that all requirement values, whether commercial or independent, are based on AMEn. In essence AME is a step closer to NE, which is touted as being the ultimate system of energy evaluation. Measuring AME or AMEn necessarily involves a live bird bioassay during which the GE of feed eaten, and resultant excreta are measured. This bioassay is straightforward in assessing diets but becomes much more complicated in assessing individual ingredients. Ingredients are substituted for basal diets or for glucose and then the ingredient value calculated from regression analysis to 100% inclusion of the test ingredient. The greater the distance from actual inclusion (e.g. 20%) level to 100% the greater the standard error of prediction. High test inclusion levels are challenging with high-fiber or high-CP ingredients and especially so for fat where maximum inclusion rarely gets above 15% of any test diet. For ingredients used at exceptionally low inclusion levels, such as amino acids, a bioassay becomes virtually impossible and so we rely on predictive estimates (van Milgen et al, 2020). Since energy comes from organic components there is potential for estimation via NIRA. Valdez and Leeson (1992a,b,c) clearly showed that NIRA has potential to predict AMEn of both diets and ingredients and was especially useful for assessing fats and oils. However, as in any NIRA measurement, prediction is reliant on robust calibration that includes calibration samples covering the entire range of anticipated AMEn.

The final potential evaluation system to consider is Net Energy. When digested energy is utilized there will be varying degrees of inefficiency associated with “rearrangement” of components necessary for ultimate deposition as fat or protein in eggs or the body of meat birds. There will also be inefficiency from a production viewpoint in that energy will be used for non-productive purposes, generally termed maintenance; with adult broiler breeders for example, the overwhelming use of energy is for maintenance. Of most economic significance is the proportion of energy deposited as protein (Nep) or fat (Nef). It is exceptionally difficult to measure NE and to date there is virtually no published information on the direct assessment of NE of ingredients for poultry. A bioassay involves first measuring AMEn and subsequently either energy deposition as fat and protein measured by measuring egg and/or carcass (body) fat and protein. Alternatively, one can estimate maintenance energy from measurement of RQ in a respiration chamber to yield NE (but not Nep and Nef). Measuring NE of diets is complicated and measuring NE of individual ingredients is a very daunting challenge. Consequently, most research on NE involves using various regression or prediction equations based on “digestible” nutrient content etc., a concept that we know does not work very accurately for AMEn. A further thought is that the NE of an ingredient or diet is as much influenced by the skill of the nutritionist during formulation, as it is the birds use of diet energy.
Energy requirements
Regardless of how energy requirements are calculated, the nutritionist must set specific energy levels during formulation. This situation is somewhat confounded with the bird’s ability to alter feed intake to regulate energy intake. Table 2 shows the ability of both layers and broilers to adjust feed intake in response to diet energy concentration.
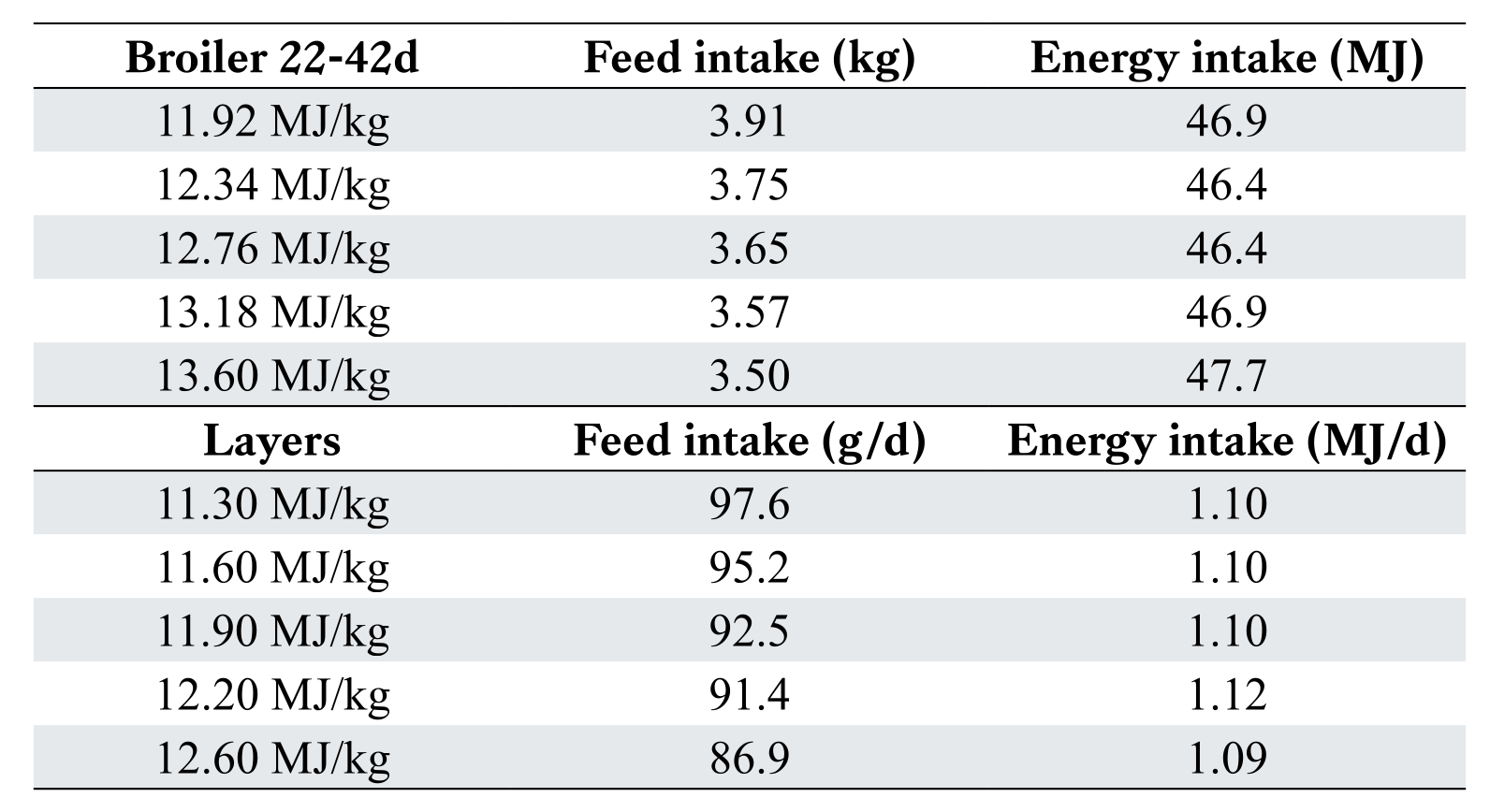
Although broilers have voracious appetites, they are still able to alter feed intake in response to a vast range of diet energy (assuming they can eat ad libitum). The almost perfect adjustment of feed intake in caged white-egg layers can be used to advantage in monitoring diet energy concentration, assuming there is accurate measurement of daily feed intake. Energy concentration is therefore more of an economic decision rather than an attempt to meet specific goals of performance. Because energy concentration impacts feed intake, then this value dictates intake of all other nutrients. The concept of maintaining an energy: protein ratio still applies to modern birds, albeit more likely replaced by consideration of energy: dig lysine. For broilers, increasing energy concentration in diets adequate in AAs invariably results in improved feed efficiency. However, adding energy to broiler diets deficient in protein/AA (even if balanced) usually has the opposite effect (Aftab, 2019).
Although there are various equations used to predict energy requirements, they invariably fail due to adequately estimate underlying feed intake. In broilers, feed intake is affected by the complex interaction involving stocking density, environmental temperature, pellet quality/size and lighting program. The situation is perhaps less complicated for layers and especially broiler breeders (Teofilo et al., 2023) where we control feed intake. Table 3 shows expected energy intake of broilers, layers and breeders.
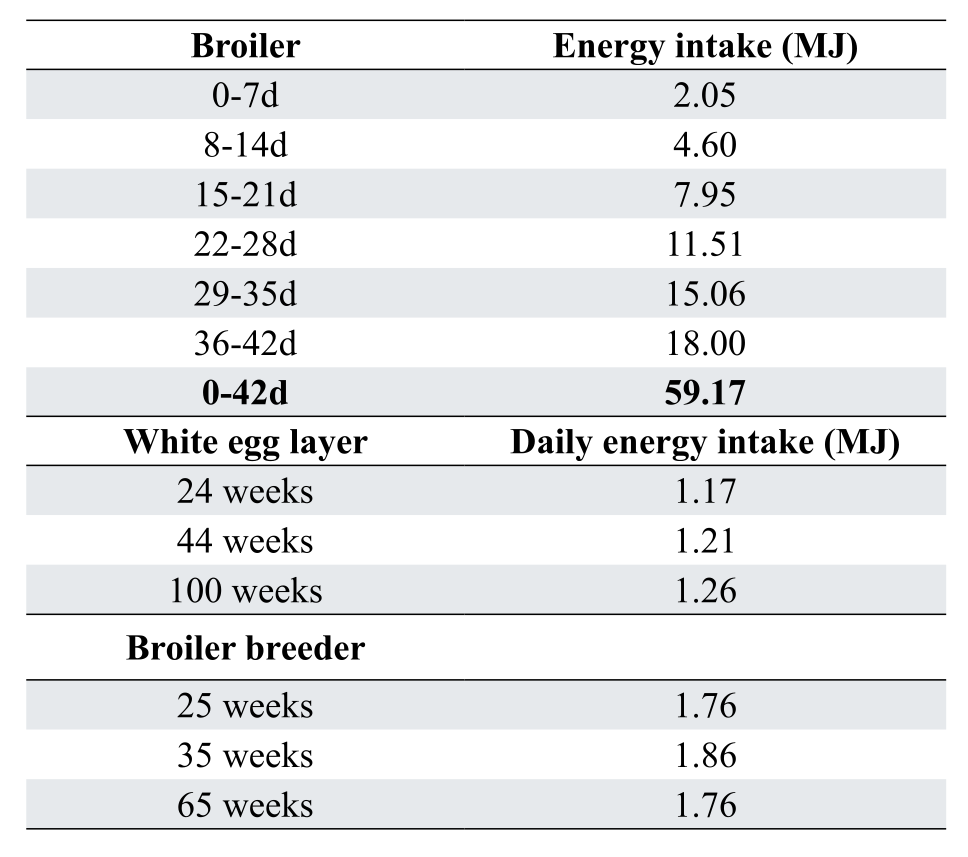
The broiler is now slightly more efficient than is the laying hen in terms of energy input per unit of “commercial” gain- for a 3 kg broiler at 42d, efficiency is around 18.8 kJ AMEn/g bodyweight gain, while for layers at peak egg mass around 44 weeks of age, a 59.5 g daily egg mass converts to around 20.5 kJ AMEn/g egg. This change has undoubtedly been caused by the propensity of modern broilers to deposit more muscle (at just over 4 MJ/kg) and less fat (at around 37 MJ/kg).
Energy requirements will obviously be impacted by environmental temperature. At temperatures much below 26 °C the bird will eat more feed to maintain body temperature. Conversely, they will eat less at higher temperatures, assuming that they are not panting which is a very energy demanding process. Measuring house temperature at first glance seems to be a straightforward task. Thermometers or temperature probes can be positioned at bird height and records collected daily. However, there is usually considerable fluctuation in temperature throughout the day. Without exact control on house temperature, how do we reconcile this temperature fluctuation in trying to calculate changes in maintenance energy and feed needs. A good example occurs with broiler breeders, since we control feed intake and so we can accommodate fluctuating environmental conditions as they occur. The traditional approach has been to simply take an average of all readings or the average of the high and low daily temperatures e.g. (26 °C+14 °C)/2 = 20 °C. However, breeders do not behave in a similar manner during the day compared with nighttime darkness. During the day most breeders are rarely in contact with other birds and so the air around them is at a temperature very similar to that recorded on the thermometer. When lights are switched off however, birds invariably sit down and are usually huddled close to their flock-mates. Sitting, rather than standing, will reduce heat loss of the bird, while huddling as a group has a great insulating effect. This behavioural change in the bird has the effect of lessening the impact of the cooler night temperatures. Simply averaging high and low temperatures, to calculate feed need, may therefore be inaccurate. Leeson and Summers (2005) proposed calculation of Effective Temperature for use in calculating energy requirements.
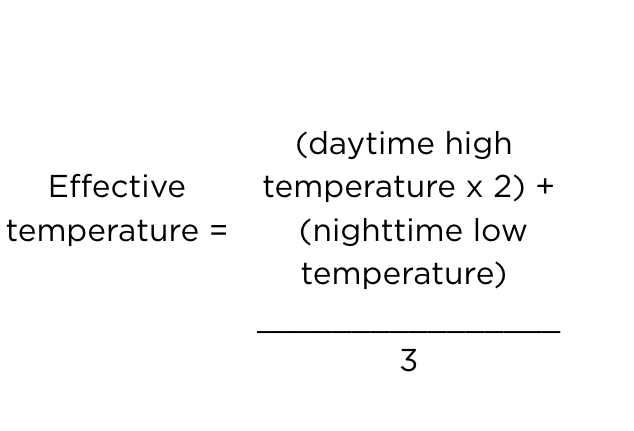
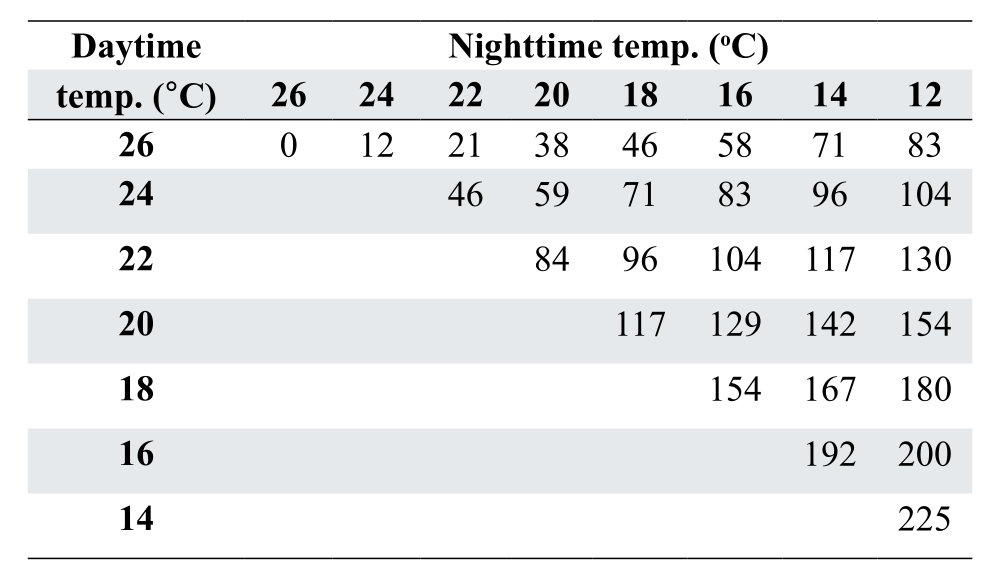
For the example detailed above, Effective Temperature becomes 24 °C rather than 20 °C. Table 4 uses this concept and describes increases in daily energy allowance needed for adult breeders maintained at various temperatures. This same concept can be used to balance energy requirements for any age or class of bird.
Energy alternatives
There are obviously no alternatives to provision of diet AMEn from the vast array of ingredients now available to the industry. The use of exogenous enzymes helps to improve DE, and formulation is designed to optimize nitrogen utilization. We can however consider various options for improving energy utilization by reducing waste in terms of Heat Increment and/or overall maintenance energy use. Figure 2 is a schematic of energy use by layers and broilers with emphasis on FMR (Fasting Metabolic Rate) maintenance needs.
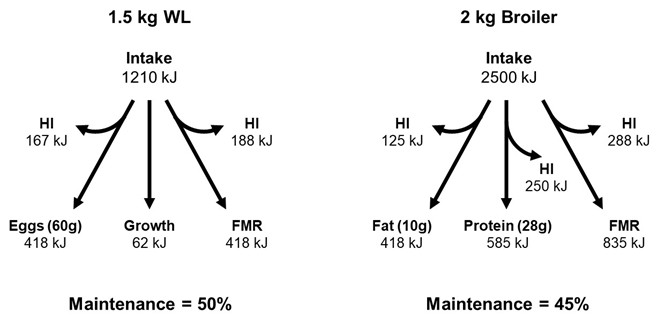
Regardless of high sustained output of our commercial strains today, we still have at least 45-50% of AMEn intake being used for “non-productive” purposes in terms of maintenance energy needs. Maintenance is comprised of the essential Fasting Metabolic Rate plus activity, plus inefficiency associated with nutrient utilization. Table 4 previously described the energetic inefficiency related to raising birds outside the thermal neutral zone, and long-term energy use will decline as globally most countries embrace controlled environment housing systems. Non-cage systems for layers (and now broilers?) obviously impose an inefficiency in energy utilization associated with variable loss in production and greater maintenance need for activity. Hendrix (2022) proposes an increase in energy requirement of at least 10% for free-range vs caged layers. For the Australian layer industry, which embraces free-range production of layers, this represents an additional $17.5 million AUD of feed energy, which seems incongruous with current interest and quantification re sustainability.
So called Heat Increment relates to the inefficiency associated with “rearrangement” of nutrients by the liver in converting from a profile provided by the feed to that required for production of muscle, adipose tissue and eggs. Rearrangement of proteins is most costly in energy need, with fats being the most efficiently utilized. In this regard, reducing crude protein, while sustaining DAA levels will always increase NE capture from the diet. However, this approach is not always economical as well as metabolically inefficient, often due to failure to balance DEB and the fact that large quantities of free synthetic amino acids may not be used with 100% efficiency, likely associated with quantity vs time of “imbalanced” supply arriving at the liver.
For broilers the reason for using pelleted diets is to reduce maintenance energy cost associated with feeding activity. The classic work of Leo Jensen at the University of Georgia showed that improvement in feed efficiency resulting from pelleting feed was not due to “improved” digestibility of heated starch, but rather an 18% reduction in the time spent feeding. Each 1% improvement in pellet quality (% of intact pellets in the feed) is equivalent to +13 kJ/kg of AMEn. Today, pelleted diets are most often used for broilers, although we often underestimate the ability of birds to consume large pellets at younger ages. Obviously the larger the pellet, the less time spent feeding. Table 5 shows the relative number of pellets consumed by broilers as pellet diameter and length are increased.
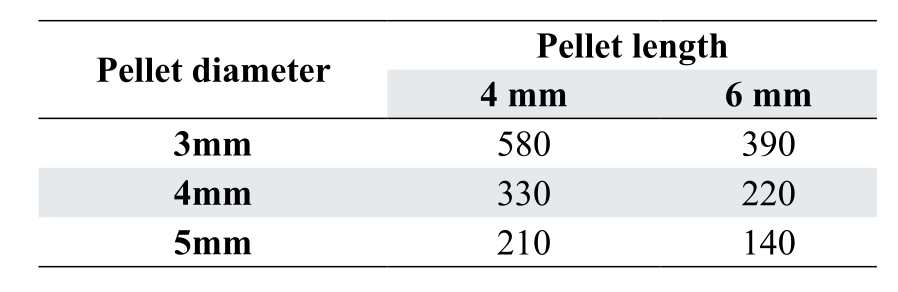
The same concept applies to layers and can be important where feed intake is compromised due to heat distress. In general, we need to limit the impact of maintenance to direct diet energy to productive purposes.
References are available on request.
From the proceedings of the Australian Poultry Science Symposium 2025, by courtesy of the Professor Steve Leeson.














